Opprinnelsen til kroppsdannelse
Fra 'Darwin's doubt' (Kap. 13)
Fra 'Darwin's doubt' (Kap. 13)
 I 1995 ble Nobel prisen i medisin gitt til to tyske forskere (Wieshaus og Nüsslein-Vollhard) for deres nye og grundige arbeid med å kartlegge genomet til bananfluer. De hadde i årtier arbeidet med mutasjonseksperimenter på disse, med nedslående resultater. De hadde nok framkalt forandringer i form av ekstremiteter, dupliseringer og endog noe funksjonsendring. Men ingenting hadde fungert i helheten. Riktignok var balanseorganer blitt endret til vinger, bare med den følge at flua mistet styringen.. Problemet var i følge Wieshaus at de trodde de hadde funnet alle genene nødvendig for å endre krop
I 1995 ble Nobel prisen i medisin gitt til to tyske forskere (Wieshaus og Nüsslein-Vollhard) for deres nye og grundige arbeid med å kartlegge genomet til bananfluer. De hadde i årtier arbeidet med mutasjonseksperimenter på disse, med nedslående resultater. De hadde nok framkalt forandringer i form av ekstremiteter, dupliseringer og endog noe funksjonsendring. Men ingenting hadde fungert i helheten. Riktignok var balanseorganer blitt endret til vinger, bare med den følge at flua mistet styringen.. Problemet var i følge Wieshaus at de trodde de hadde funnet alle genene nødvendig for å endre krop psplanen til Drosophila (typen bananflue). Likevel er ikke resultatene lovende som råmateriale for makro-evolusjon. Det neste spørsmålet er hva som ville være de rette mutasjoner for makro-evolusjon. Og vi vet ikke svaret på det..
psplanen til Drosophila (typen bananflue). Likevel er ikke resultatene lovende som råmateriale for makro-evolusjon. Det neste spørsmålet er hva som ville være de rette mutasjoner for makro-evolusjon. Og vi vet ikke svaret på det..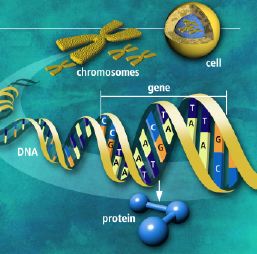 Utviklingsbiologien kaller prosessen der embryoet utvikler seg til modne organismer (gjelder flercellede organismer) for ontogeni. Feltet har stadig økt vår forståelse for hvordan kroppsplaner oppstår. Mye av kunnskapen har kommet gjennom såkalte modell-systemer, organismer som biologer lett kan mutere i laben (bananfluen Drosophila, samt en rundorm etc.) Selv om det er forvirrende ulikheter, er det ett felles trekk med all ontogeni: Den starter med ei celle og slutter med mange ulike celler. I de fleste arter starter det med ett befruktet egg. Med en gang egget deler seg i dets datter-celler, blir egget et embryo. Da begynner det å stile mot et vel-definert mål, nemlig en voksen form som kan reprodusere. For å nå det målet må embryoet produsere mange spesialiserte celle-typer på korrekt sted og tid.
Utviklingsbiologien kaller prosessen der embryoet utvikler seg til modne organismer (gjelder flercellede organismer) for ontogeni. Feltet har stadig økt vår forståelse for hvordan kroppsplaner oppstår. Mye av kunnskapen har kommet gjennom såkalte modell-systemer, organismer som biologer lett kan mutere i laben (bananfluen Drosophila, samt en rundorm etc.) Selv om det er forvirrende ulikheter, er det ett felles trekk med all ontogeni: Den starter med ei celle og slutter med mange ulike celler. I de fleste arter starter det med ett befruktet egg. Med en gang egget deler seg i dets datter-celler, blir egget et embryo. Da begynner det å stile mot et vel-definert mål, nemlig en voksen form som kan reprodusere. For å nå det målet må embryoet produsere mange spesialiserte celle-typer på korrekt sted og tid. Under ontogenien dobles antall celler hele tiden, ofte i en rasende fart. Antall celle-delinger og totalt antall celler reflekterer tallet på ulike celle-typer den modne organismen trenger. Dette innebærer igjen å produsere ulike proteiner for ulike celle-typer. Hvert protein må differensieres, fordi de gjerne utfører helt ulike oppgaver. Under ontogenien må da de rette genene bli slått av/på for å sikre at rett protein blir produsert til rett tid i de rette celle-typer. Spesifikke proteiner spiller aktive roller i regulering hvordan gener framtrer for å bygge andre proteiner. Proteinene som spiller denne rollen er kjent som transkripsjons-regulatorer(TRs) eller transkripsjons-faktorer (TFs). Disse knytter seg direkte til spesifikke steder i DNA og enten hindrer eller aktiver transkripsjon av spesifikke gener inn i RNA. De inneholder instruksjoner om hvilke gener de skal slå av eller på. Den tredimensjonale geometrien utøver karakteristiske trekk ved DNA-binding: inkl. et område av 61 aminosyrer som pakker seg omkring den doble DNA-spiralen.
Under ontogenien dobles antall celler hele tiden, ofte i en rasende fart. Antall celle-delinger og totalt antall celler reflekterer tallet på ulike celle-typer den modne organismen trenger. Dette innebærer igjen å produsere ulike proteiner for ulike celle-typer. Hvert protein må differensieres, fordi de gjerne utfører helt ulike oppgaver. Under ontogenien må da de rette genene bli slått av/på for å sikre at rett protein blir produsert til rett tid i de rette celle-typer. Spesifikke proteiner spiller aktive roller i regulering hvordan gener framtrer for å bygge andre proteiner. Proteinene som spiller denne rollen er kjent som transkripsjons-regulatorer(TRs) eller transkripsjons-faktorer (TFs). Disse knytter seg direkte til spesifikke steder i DNA og enten hindrer eller aktiver transkripsjon av spesifikke gener inn i RNA. De inneholder instruksjoner om hvilke gener de skal slå av eller på. Den tredimensjonale geometrien utøver karakteristiske trekk ved DNA-binding: inkl. et område av 61 aminosyrer som pakker seg omkring den doble DNA-spiralen.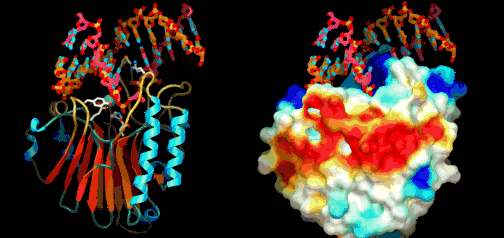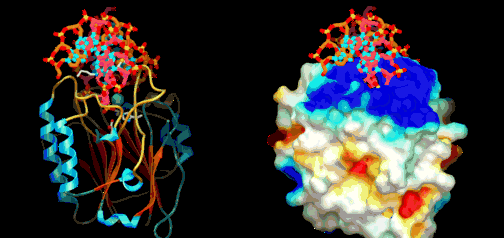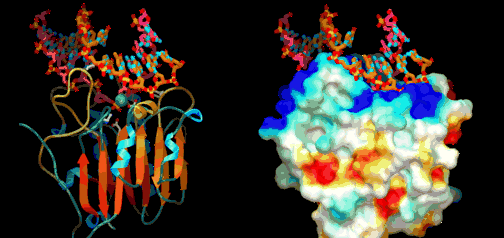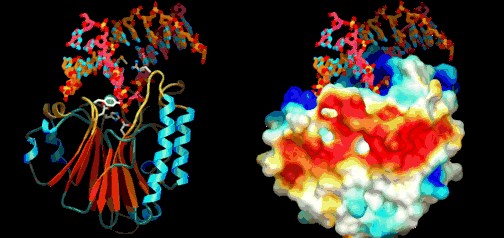 Eric Davidson er den biologen som mest dyptgående har undersøkt regulerings-logikken i dyrs utvikling. Davidson observerte at cellene til et individuelt dyr, samme hvor variert i form eller funksjon, generelt besto av identiske genomer. Gjennom livssyklusen til en organisme, så uttrykte genomet til disse spesialiserte cellene bare en liten del av sitt DNA til en gitt tid, og produserte dermed ulikt RNA som resultat. Dette faktum antyder sterkt at det finnes ett genetisk kontrollsystem for hele organismen, som skrur brytere på og av etter hvor en er i embryo-utviklingen. Likeså at et slikt system styrer utviklingen fra egg til moden organisme, i det ulike celle typer dannes. Britten og Davidson la fram en teori ang. dette i 1969. Selv om lite da var kjent, deduserte de at et slikt system måtte være virksomt. Det var for det første fordi hundrevis av spesialiserte celler oppsto gjennom utviklingen av dyr, fra embryo til fullt utviklet. For det andre at hver celle inneholdt samme genom, så sluttet de at et slags kontroll-system måtte bestemme hvilke gener som uttrykkes i ulike celler til ulike tider, for å sikre differensiering av ulike celletyper fra hverandre. Derfor måtte det være en regulerende logikk som overstyrte og koordinerte, hvordan genomet uttrykte seg til enhver tid.
Eric Davidson er den biologen som mest dyptgående har undersøkt regulerings-logikken i dyrs utvikling. Davidson observerte at cellene til et individuelt dyr, samme hvor variert i form eller funksjon, generelt besto av identiske genomer. Gjennom livssyklusen til en organisme, så uttrykte genomet til disse spesialiserte cellene bare en liten del av sitt DNA til en gitt tid, og produserte dermed ulikt RNA som resultat. Dette faktum antyder sterkt at det finnes ett genetisk kontrollsystem for hele organismen, som skrur brytere på og av etter hvor en er i embryo-utviklingen. Likeså at et slikt system styrer utviklingen fra egg til moden organisme, i det ulike celle typer dannes. Britten og Davidson la fram en teori ang. dette i 1969. Selv om lite da var kjent, deduserte de at et slikt system måtte være virksomt. Det var for det første fordi hundrevis av spesialiserte celler oppsto gjennom utviklingen av dyr, fra embryo til fullt utviklet. For det andre at hver celle inneholdt samme genom, så sluttet de at et slags kontroll-system måtte bestemme hvilke gener som uttrykkes i ulike celler til ulike tider, for å sikre differensiering av ulike celletyper fra hverandre. Derfor måtte det være en regulerende logikk som overstyrte og koordinerte, hvordan genomet uttrykte seg til enhver tid.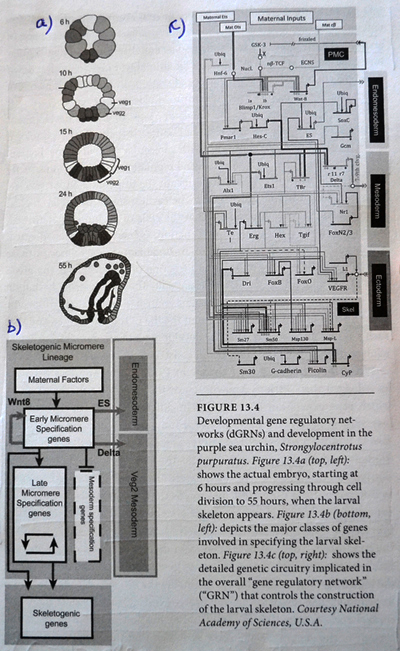 Den fantastiske kompleksiteten de fant framstilles på figure 13.4 her ved siden. Del a viser utvikling av embryoet i en alder fra 6 til 55 timer. Øverst er det 4 runder med celledeling og 16 celler (2 opphøyd i 4). I de neste fire stadiene ser vi en tiltagende spesialisering i kroppsformen. Del b er et skjematisk diagram over hovedklassene av gener og cellevev, i løpet av embryo-utviklingen. Sammenhengen markeres med kontrollpiler. Fig 13.c viser 'det genetiske kretskortet' (i følge Davidson) som skrur på de spesifikke genene, som igjen produserer det nødvendige strukturelle proteinet for å danne sjøpiggsvinet.
Den fantastiske kompleksiteten de fant framstilles på figure 13.4 her ved siden. Del a viser utvikling av embryoet i en alder fra 6 til 55 timer. Øverst er det 4 runder med celledeling og 16 celler (2 opphøyd i 4). I de neste fire stadiene ser vi en tiltagende spesialisering i kroppsformen. Del b er et skjematisk diagram over hovedklassene av gener og cellevev, i løpet av embryo-utviklingen. Sammenhengen markeres med kontrollpiler. Fig 13.c viser 'det genetiske kretskortet' (i følge Davidson) som skrur på de spesifikke genene, som igjen produserer det nødvendige strukturelle proteinet for å danne sjøpiggsvinet. Developmental gen regulatory networks (Dgrns) motstår mutasjonsendringer fordi de er hierarkisk organisert. Det innebærer at noen Dgrns kontrollerer andre Dgrns. I senteret av dette regulerende hierarkiet, er regulerende nettverk som spesifiserer akse og helhetlig kroppsplan under utviklingen. Selv små endringer i disse Dgrns vil medføre katastrofale virkninger på organismen. Likevel er det nettopp det som trengs om akse og global form på dyret skal variere: Nettopp de kretsløpene som ikke kan endres uten ødeleggelse som følge, må endre seg, om kroppsformen skal endres.
Developmental gen regulatory networks (Dgrns) motstår mutasjonsendringer fordi de er hierarkisk organisert. Det innebærer at noen Dgrns kontrollerer andre Dgrns. I senteret av dette regulerende hierarkiet, er regulerende nettverk som spesifiserer akse og helhetlig kroppsplan under utviklingen. Selv små endringer i disse Dgrns vil medføre katastrofale virkninger på organismen. Likevel er det nettopp det som trengs om akse og global form på dyret skal variere: Nettopp de kretsløpene som ikke kan endres uten ødeleggelse som følge, må endre seg, om kroppsformen skal endres.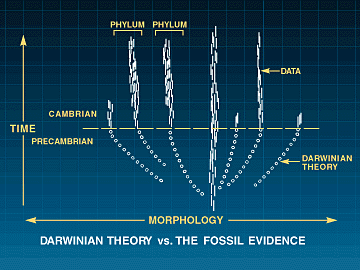 OG nye dGRNs. OM ingen av disse kan endres ved mutasjoner, uten å ødelegge eksisterende utviklingsprogram (og dermed dyreliv), så vil ikke naturlig utvalg ha noe å favorisere. Dermed vil utviklingen av nye dyreformer holde opp på det stadiet. Darwins tvil om fossilfunn i Cambrium er forsterket: Fossilene mangler, men i tillegg har vi et dyptgående problem med konstruksjons begrensinger. Problemet 'å bygge en ny form for dyreliv, ved gradvis å endre et tett integrert system av genetiske komponenter og deres produkter infiltrert i hverandre'. Ennå gjenstår mer formidable problemer for neo-Darwinismen:
OG nye dGRNs. OM ingen av disse kan endres ved mutasjoner, uten å ødelegge eksisterende utviklingsprogram (og dermed dyreliv), så vil ikke naturlig utvalg ha noe å favorisere. Dermed vil utviklingen av nye dyreformer holde opp på det stadiet. Darwins tvil om fossilfunn i Cambrium er forsterket: Fossilene mangler, men i tillegg har vi et dyptgående problem med konstruksjons begrensinger. Problemet 'å bygge en ny form for dyreliv, ved gradvis å endre et tett integrert system av genetiske komponenter og deres produkter infiltrert i hverandre'. Ennå gjenstår mer formidable problemer for neo-Darwinismen:Fra Darwins Doubt kap. 14

 Eukaryote celler har interne skjelett for å gi dem form og stabilitet. Disse skjelettene har ulike slag av fibre, inkludert mikrotuber . Strukturen og lokalisering av mikrotubene i cytoskjelettet influerer formen på og utviklingen av embryoet. Oppstillingen av mikrotuber innen embryo-celler, hjelper å distribuere essentielle proteiner som brukes under utvikling til spesifikke lokasjoner i cellen. Når de er dannet, utfører slike proteiner funksjoner kritiske for utviklingen av embryoet. Forutsetningen er at de blir levert til sine korrekte plasseringer, ved hjelp av preeksistente, presist formulerte oppstillinger av mikrotuber og cytoskjelett. Slik blir den presise arrangeringen av mikrotuber i cytoskjelettet en form for kritisk, strukturert informasjon.
Eukaryote celler har interne skjelett for å gi dem form og stabilitet. Disse skjelettene har ulike slag av fibre, inkludert mikrotuber . Strukturen og lokalisering av mikrotubene i cytoskjelettet influerer formen på og utviklingen av embryoet. Oppstillingen av mikrotuber innen embryo-celler, hjelper å distribuere essentielle proteiner som brukes under utvikling til spesifikke lokasjoner i cellen. Når de er dannet, utfører slike proteiner funksjoner kritiske for utviklingen av embryoet. Forutsetningen er at de blir levert til sine korrekte plasseringer, ved hjelp av preeksistente, presist formulerte oppstillinger av mikrotuber og cytoskjelett. Slik blir den presise arrangeringen av mikrotuber i cytoskjelettet en form for kritisk, strukturert informasjon. Fra Origo 141-om vitenskap, skapelse og etikk
 Utviklingsbiologer oppdager flere og flere måter der avgjørende informasjon kommer fra fostercellens form og struktur. Informasjonen som benyttes for å bygge kroppsstrukturen kommer både fra det ubefruktede og befruktede egg. Disse informasjonskildene betegner forskerne som epigenetiske ('over det genetiske'). Helt siden 1980 årene har mange forskere ment at nødvendig informasjon til komplekse biologiske systemer overgår den som ligger i DNAet. Når proteiner er framstilt skal de arrangeres i overordnede strukturer. Om ikke, blir ikke nødvendige organer og vev dannet. I følge St. C. Meyer er fostercellens tredimensjonale struktur en viktig rolle under fosterutviklingen. Her har biologer funnet ulike kilder til epigenetisk informasjon: Cytoskjelettets organisering, membranens oppbygning (målområder og ionekanaler) samt sukkerkoden.
Utviklingsbiologer oppdager flere og flere måter der avgjørende informasjon kommer fra fostercellens form og struktur. Informasjonen som benyttes for å bygge kroppsstrukturen kommer både fra det ubefruktede og befruktede egg. Disse informasjonskildene betegner forskerne som epigenetiske ('over det genetiske'). Helt siden 1980 årene har mange forskere ment at nødvendig informasjon til komplekse biologiske systemer overgår den som ligger i DNAet. Når proteiner er framstilt skal de arrangeres i overordnede strukturer. Om ikke, blir ikke nødvendige organer og vev dannet. I følge St. C. Meyer er fostercellens tredimensjonale struktur en viktig rolle under fosterutviklingen. Her har biologer funnet ulike kilder til epigenetisk informasjon: Cytoskjelettets organisering, membranens oppbygning (målområder og ionekanaler) samt sukkerkoden. Målområder i membranen
Målområder i membranen Sukkerkoden
Sukkerkoden alltid ført til skadelige eller katastrofale konsekvenser. Eksperimenter har gjennomgående vist at ved omfattende endringer i epigenetisk informasjon, så har ikke embryoet en sjanse til å overleve i naturen, enn si å formere seg.
alltid ført til skadelige eller katastrofale konsekvenser. Eksperimenter har gjennomgående vist at ved omfattende endringer i epigenetisk informasjon, så har ikke embryoet en sjanse til å overleve i naturen, enn si å formere seg.
| Mønstre i cytoskjelettet | Plassering avgjør- en form for kritisk informasjon | Arves direkte |
| Mikrotubuler | Lokasjon og arrangement avgjør arv | Testbart |
| Ione-kanaler og elektromagnetiske felt | Arrangering og plassering av ione-kanaler kan påvirke dyrets utvikling | Kunstige felt kan forårsake celle-vandring. |
| Sukkermolekyler på cellemembran | Plassering spiller kritisk rolle i funksjonen de har |
Testet på |
| Mål-punkter på celle-membran | Spesifikk lokasjon og fordeling avgjør funksjon | Plassering avgjøres ikke av genprodukt alene |
| Folding av proteiner | Mange proteiner folder seg ulikt avhengig av omgivelsene | Påvist i sammenlignende utviklingsbiologi |
| Morfogenetiske proteiner, regulerende gener og dGRNs | Disse avhenger alle av plasseringen av spesifikke, informasjonsrike, tidligere cellestrukturer | En har aldri opplevd at dGRNs er blitt endret med livskraftig avkom som følge |
| Cellestruktur | Resultat av både genetikk og tidligere 3-dim-struktur og organisering i celle, cellemembran, og cytoskjelett | Påvist ved forsøk at snitt i cellemembran arves direkte på encellede organismer |
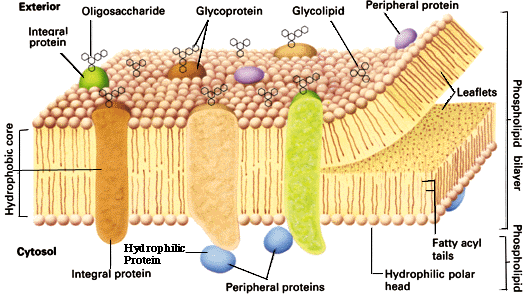 Cellemembranens målområder spiller en avgjørende rolle i utviklingen av embryoet, ved å tiltrekke molekyler til spesifikke plasser på indre celleoverflate. Når mange proteiner folder seg ulikt etter hvor de befinner seg i cellulær kontekst, så viser det kontekst-avhengig informasjon.
Cellemembranens målområder spiller en avgjørende rolle i utviklingen av embryoet, ved å tiltrekke molekyler til spesifikke plasser på indre celleoverflate. Når mange proteiner folder seg ulikt etter hvor de befinner seg i cellulær kontekst, så viser det kontekst-avhengig informasjon.
 2. I den grad cellestrukturen er utsatt for endringer, er disse endringene i all hovedsak sannsynlige, å ha skadelige eller katastrofale konsekvenser. Sperman and Mangold utførte et eksperiment der de påtvang endringer i et viktig lager for epigenetisk informasjon, i løpet av embryo-perioden. Selv om resultatet hadde et interessant utseende, hadde det ikke sjanse til å leve opp, langt mindre formere seg.Darwins bok (Origin of Species) skulle f.o.f. være en forklaring på oppkomsten av nye arter . Han vedkjente at mønsteret for tilsynekomsten av dyreartene ikke skjedde i henhold til hans gradvise utviklings-bilde for livets historie. Mens han levde ble det ansett være et spørsmål om ufullstendighet i fossilmateriale. Problemet har ikke kommet nærmere noen løsning, men nå er det et større og mer funamentalt problem som påvirker hele den darwinistiske konstruksjonen: Neo-darwinstisk metodologi kan verken gjøre rede for opprinnelsen til nødvendig genetisk eller epigenetisk informasjon for å produsere nye livsformer.
2. I den grad cellestrukturen er utsatt for endringer, er disse endringene i all hovedsak sannsynlige, å ha skadelige eller katastrofale konsekvenser. Sperman and Mangold utførte et eksperiment der de påtvang endringer i et viktig lager for epigenetisk informasjon, i løpet av embryo-perioden. Selv om resultatet hadde et interessant utseende, hadde det ikke sjanse til å leve opp, langt mindre formere seg.Darwins bok (Origin of Species) skulle f.o.f. være en forklaring på oppkomsten av nye arter . Han vedkjente at mønsteret for tilsynekomsten av dyreartene ikke skjedde i henhold til hans gradvise utviklings-bilde for livets historie. Mens han levde ble det ansett være et spørsmål om ufullstendighet i fossilmateriale. Problemet har ikke kommet nærmere noen løsning, men nå er det et større og mer funamentalt problem som påvirker hele den darwinistiske konstruksjonen: Neo-darwinstisk metodologi kan verken gjøre rede for opprinnelsen til nødvendig genetisk eller epigenetisk informasjon for å produsere nye livsformer.